Immunological properties of gene vaccines delivered by different routes
S.C. Oliveira1, G.M.S. Rosinha1, C.F.A. de-Brito1, C.T. Fonseca1, R.R. Afonso1, M.C.M.S. Costa1, A.M. Goes1, E.L. Rech3 and V. Azevedo2
Departamentos de 1Bioquímica e Imunologia and 2Biologia Geral,
Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brasil
3Centro Nacional de Pesquisas de Recursos Genéticos e Biotecnologia, EMBRAPA,
Brasília, DF, Brasil
![]()
Gene vaccines represent a new and promising approach to control infectious diseases, inducing a protective immune response in the appropriate host. Several routes and methods of genetic immunization have been shown to induce antibody production as well as T helper (Th) cell and cytotoxic T lymphocyte activation. However, few studies have compared the nature of the immune responses generated by different gene vaccination delivery systems. In the present study we reviewed some aspects of immunity induced by gene immunization and compared the immune responses produced by intramuscular (im) DNA injection to gene gun-mediated DNA transfer into the skin of BALB/c mice. Using a reporter gene coding for ß-galactosidase, we have demonstrated that im injection raised a predominantly Th1 response with mostly IgG2a anti-ßgal produced, while gene gun immunization induced a mixed Th1/Th2 profile with a balanced production of IgG2a and IgG1 subclasses. Distinct types of immune responses were generated by different methods of gene delivery. These findings have important implications for genetic vaccine design. Firstly, a combination between these two systems may create optimal conditions for the induction of a broad-based immune response. Alternatively, a particular gene vaccine delivery method might be used according to the immune response required for host protection. Here, we describe the characteristics of the immune response induced by gene vaccination and the properties of DNA involved in this process.
Key words: gene vaccination, naked DNA, gene gun, cytokines, immune response
Introduction
Gene vaccination is a novel approach to immunize animals and humans against pathogenic microorganisms. Introduction of nucleic acid coding for a specific antigen into a living cell results in immune activation of the host against the gene-delivered immunogen and in some cases protection (1). The use of a DNA plasmid carrying vaccine gene antigens is a great advantage compared to the unwanted effects of the introduction of a pathogen or recombinant protein lacking conformational epitopes. Therefore, genetic immunization is by far the most powerful technique, which resembles a natural infection without side effects.
The production of genetic vaccines in host cells using DNA has a number of immunological features important for the success of vaccination. There are some unique properties of DNA vaccines which represent advantages over other immunization procedures: i) no risk of infection, ii) inducing a long-lived immune response, iii) eliciting humoral and cell-mediated immunity, iv) facilitating the use of polyvalent vaccines, v) good stability at low and high temperatures, vi) increasing cytolytic T-cell responses, vii) easy preparation and purification, and viii) low cost.
Historically, Wolff et al. (2) in 1990 demonstrated that nonreplicating DNA plasmids encoding reporter genes could express encoded proteins in muscle cells following DNA injection. Tang and colleagues (3) in 1992 reported that using biolistic-mediated gene transfer by gene gun inoculation, they could immunize animals with plasmid DNA coated on gold beads. And in 1993, the Merck research group (1) induced protective immunity in mice by immunization with DNA encoding human influenza virus A antigen. Therefore, immune responses have currently been induced by injection of naked DNA in saline, or by biolistic-mediated gene transfer using a gene gun (4).
In the battle against infectious diseases, immune responses to antigens encoded by and delivered as plasmid DNAs have been raised in a variety of species (humans, mice, cattle, dog, ferrets, nonhuman primates) against several different immunogens from pathogens such as influenza virus (1), bovine herpesvirus (5), human immunodeficiency virus-1 (6), hepatitis B (7), malaria (8), and tuberculosis (9), among others. Besides permitting a specific immune response targeted to the encoded antigen, plasmid DNA may be used to modulate the immune system in different ways. First, the DNA molecule has its own immunostimulatory properties, functioning as a natural adjuvant. Alternatively the introduction of plasmid DNA encoding a particular cytokine gene or costimulatory molecules may bias the immune response activating the host immune system.
Successful vaccination depends on the induction of an appropriate immune response capable of protecting the host against disease. Here, we are going to discuss the qualitative and quantitative differences in the immune response elicited during gene vaccination by intramuscular (im) injection or particle bombardment using a gene gun. Both systems were able to raise a strong humoral response; however, total IgG levels following gene gun immunization were a little higher compared to im injection. Regarding the cytokine profile, our data suggest that im injection elicited a preferential Th1 type of immune response and gene gun vaccination elicited a mixed Th1/Th2 profile. Therefore, it is important to stress that, according to the pattern of immune response required to achieve protection, a specific gene delivery system might be used.
Gene vaccine delivery systems
Currently, there are two main delivery systems used in genetic immunization: plasmid DNA injected intramuscularly or DNA coated onto gold beads and transferred into the epidermis or dermis by a biolistic process (gene gun).
Intramuscular injection
This is the most widely used method for immunization and it consists of direct injection of naked DNA into skeletal muscle. Plasmid DNA in some instances is injected into muscle directly in saline solution (1), or after injection of toxins or a local anesthetic (e.g. bupivacaine) to cause necrosis and regeneration of the injected muscle, increasing the expression of the encoded antigen, and therefore amplifying the immunological response (10). In our laboratory, we injected 50 µl of 10 mM cardiotoxin (snake venom) into the quadriceps muscles of each mouse five days prior to gene inoculation. However, it is unclear whether the enhancement of the immune response is directly mediated by the elevated expression of antigen in regenerating muscle cells, or via the uptake and expression of the gene of interest by antigen-presenting cells (APC) recruited to the site of tissue damage.
Humoral and cell-mediated immune responses have been induced by direct im injection of plasmid DNA encoding immunogens. An antibody response was first reported against an influenza virus protein in mice (1), and specific cytotoxic T cell responses were also detected in different systems such as HIV and hepatitis B following genetic immunization (11,12). Protective immunity was first demonstrated in mice injected intramuscularly with nucleoprotein DNA of influenza virus (1). In this model, researchers have indicated that both CD4+ and CD8+ T cells contributed to achieve protection. Protective immune responses have also been demonstrated in mice against Leishmania major, Plasmodium yoelii, Mycobacterium tuberculosis, dengue virus, and herpes simplex virus (8,9,13-15).
Biolistic-mediated gene transfer (gene gun)
Biolistic is a method which uses a helium gas pressure-driven device (gene gun) to deliver gold particles coated with plasmid DNA directly into the skin. When gene vaccines are administered by gene gun technology most of the plasmid DNA is taken up by keratinocytes and some dermal fibroblasts, which become transfected and produce the encoded antigen.
Humoral responses using biolistic were first demonstrated by Tang et al. (3) using plasmids encoding human growth hormone and human a-1 anti-trypsin. Subsequently, Fynan et al. (16) and Webster et al. (17) used the gene gun method in a mouse influenza virus model. Regarding cell-mediated immunity elicited by biolistic, the first evidence was reported by Hui et al. (18) who bombarded surgically exposed tissues with MHC H-2Kb molecules and detected allospecific cytotoxic responses. Cytolytic responses were also developed in mice bombarded directly into the skin with HIV gp120 and env genes (19). Protective immunity has been studied using biolistic in different infectious disease systems such as lymphocytic choriomeningitis virus (LCMV), Plasmodium berghei, Ebola virus, pseudorabies virus, rotavirus, and Mycoplasma pulmonis (20-25).
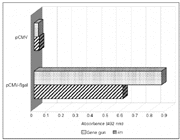
To study the immune response induced by im injection and biolistic-mediated gene transfer, we have immunized BALB/c mice with a plasmid carrying the reporter gene coding for ß-galactosidase (pCMV-ßgal). Figure 1 shows that specific anti-ß-galactosidase antibody responses were raised using both gene delivery systems; however, IgG levels were slightly higher following gene gun vaccination when compared to im injection. These data demonstrate the relative effectiveness of these two systems, the gene gun and im injection, to mount a specific humoral response. Total IgG responses can be detected as early as two weeks after a single immunization with pCMV-ßgal using either gene delivery method. However, we have shown that only after the third immunization did vaccinated mice reach a peak of total IgG production. The qualitative difference in the humoral response observed in Figure 1 between gene gun and im vaccination is probably due to several reasons. According to Barry and Johnston (26), one of them is the fact that im injection places the DNA extracellularly where most of it may be rapidly degraded by nucleases. In contrast, biolistic delivers the DNA inside the cells, bypassing this initial reduction in functional plasmids.
|
Figure 1 - Analysis of the immune responses induced in BALB/c mice following gene vaccination with pCMV-ßgal plasmid by gene gun or im injection. Mice (ten per group) were injected with cardiotoxin (10 mM) ten days prior to im vaccination with 100 µg of plasmid DNA into the quadriceps muscles. For gene gun immunization, animals were bombarded on the shaved abdomen with two shots per animal (below 0.5 µg of DNA per shot). Using both methods, mice were vaccinated three times at two-week intervals. The control group received pCMV plasmid DNA without the gene coding for ß-gal. On day 45, animals were bled and total IgG anti-ß-galactosidase were measured by indirect ELISA. Results are reported as mean values of triplicates. |
[View larger version of this image (47 K GIF file)]
Antigen expression following genetic immunization
In order to develop a proper immune response during a DNA-based immunization, gene expression of the encoded antigen must take place, followed by processing and presentation of the protein produced by APC. During muscle inoculations most of the antigen expression occurs in skeletal muscle, whereas after gene gun immunization, expression is mostly in keratinocytes. Robinson (27) proposed different mechanisms by which professional APC could acquire DNA-expressed antigens: i) transfected muscle cells or keratinocytes could produce antigen that is picked-up by APC; ii) APC could also undergo direct transfection at the target site (e.g., Langerhans cells in the dermis), or be transfected by DNA that moved from the target site through blood or lymph, to transfect APC in lymphoid tissues. Since neither muscle cells nor keratinocytes constitutively express MHC class II or the costimulatory molecules required to activate T-helper (Th) cells, their role in antigen processing and presentation needs to be better understood.
One of the most important safety considerations for the use of gene vaccines is the possibility of the integration of plasmid DNA into the genome of the host cell. Integration would produce insertional mutagenesis with the potential of activating oncogenes or inactivating tumor suppressor genes. Nichols et al. (28), in a study carried out in an attempt to detect integration of a plasmid DNA carrying the nucleoprotein gene into the mouse cell genome, failed to detect insertion of the vaccine plasmid. These investigators concluded that no integration was detected at a sensitivity of one plasmid copy per 150,000 nuclei, which indicates that any mutation frequency induced by gene vaccination would be approximately three orders of magnitude lower than the spontaneous mutation frequency.
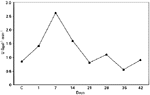
In Figure 2, we show ß-galactosidase activity in mouse cells after gene gun immunization with pCMV-ßgal plasmid DNA. ß-Galactosidase gene expression peaked at one week following gene inoculation and decreased to basal levels during the third week after vaccination. These data demonstrate that mouse cells were transiently transfected by particle bombardment, corroborating the fact that plasmid DNA-based vaccines are designed to remain nonintegrated in host cells. Nevertheless, the immune response obtained with gene vaccines is sustained for a long period of time, and in some cases a single im injection of DNA maintains high antibody levels for at least 17 months (29). The fact that muscle cells are postmitotic leaves the possibility of gene expression for months or even years. In contrast, gene gun or dermal immunization seems to induce a shorter duration of antigen synthesis compared to im vaccination. Overall, longevity of immune responses in DNA-based immunization is a phenomenon that still remains to be elucidated.
|
Figure 2 - Quantitative determination of ß-galactosidase gene expression following gene gun immunization with pCMV-ßgal. BALB/c mice (five per group) were vaccinated by the biolistic method with one shot per animal (below 0.5 µg of DNA) in the ear skin region. Every week after immunization ß-galactosidase activity was measured by a spectrophotometric assay using o-nitrophenol-ß-D-galactoside (ONPG). Results are reported as mean values of ß-galactosidase units (U/ßgal) over a period of time (in minutes). Units = (380 x A420) divided by time, where 380 is a constant such that one unit is equivalent to conversion of one nanomole of ONPG per minute at 37oC. C, Control. |
[View larger version of this image (6 K GIF file)]
Different patterns of immune response induced by gene vaccination
For gene immunization to be considered a viable approach to vaccination, the immune response obtained must be shown to be sufficient to protect the host against infection by a pathogen. As a matter of fact, protection was successfully achieved against different infectious disease models following genetic immunization, as reported earlier (13-15,20-25). In a well-studied virus infection system such as influenza, im injection of the nucleoprotein gene afforded protection as a result of cell-mediated immunity since passive transfer of serum from immune mice did not engender protective immunity (1). Depletion experiments demonstrated that both CD8+ and CD4+ T cell subpopulations were involved in host protection against infection.
Gene vaccines consisting of a diverse array of pathogen antigens are highly effective in inducing a strong MHC class I-restricted cytotoxic T lymphocyte (CTL) response as well as Th cell activation, and consequently antibody production. Regarding CTL immune responses, genetic immunization provides a mechanism to achieve intracellular antigen synthesis and to induce MHC class I presentation, mimicking a viral infection. In the case of CD4+ T cells, subsets are distinguished according to the cytokines produced (30). In mice, the type 1 pattern of immune response activates macrophages to kill intracellular parasites, promotes the delayed-type hypersensitivity reaction, and increases IgG2a and IgG3 isotypes via IFN-g production (30). The type 2 profile helps to produce IgG1 and IgE isotypes via IL-4 secretion and activates eosinophils via IL-5 production (31).
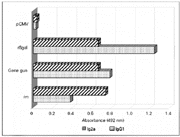
Figure 3 shows the production of IgG1 and IgG2a subclasses after DNA immunization with pCMV-ßgal by im injection or gene gun compared to inoculation with recombinant ß-galactosidase in incomplete Freund's adjuvant. These data suggest a different cytokine profile induced according to the antigen delivery system used in gene vaccination. Intramuscular injection of pCMV-ßgal plasmid DNA induces a Th1 response compared to gene gun-mediated DNA transfer which elicits a mixed Th1/Th2 pattern of cytokines based on the production of antigen-specific IgG2a and IgG1. This result agrees with studies by Leclerc et al. (32) and Tighe et al. (33) who reported that recombinant ß-galactosidase plus adjuvant induced a Th2 cytokine profile and im injection of the gene-encoded protein promoted a Th1 pattern of immune response with IFN-g production.
|
Figure 3 - Antibody profiles induced by im injection or gene gun vaccination with plasmid DNA compared to recombinant protein immunization. The IgG2a and IgG1 responses produced in BALB/c mice (five per group) were measured after gene vaccination with pCMV-ßgal or pCMV using the two gene delivery systems according to the protocol described in Figure 1. Antibody subclass responses were also detected following subcutaneous injection of 10 µg of recombinant ß-galactosidase (rßgal) per animal plus incomplete Freund's adjuvant. Mice were bled after the third immunization on day 45, and antibody levels were measured by indirect ELISA. Data are reported as mean values of triplicates. |
[View larger version of this image (44 K GIF file)]
Different immune responses produced by gene gun and im immunization are probably due to the adjuvant effect of the plasmid DNA itself. There is a critical difference between these two methods in the amount of DNA required to produce equivalent antigen expression. In our laboratory, we have used less than 0.5 µg of DNA per gene gun shot, with two shots per mice. For im injection, we delivered 100 µg of DNA into mouse quadriceps muscles per immunization. Bacterial DNA and oligonucleotides containing CpG motifs have been shown to activate B cells and to induce secretion of IL-6, IFN-g, IL-12 and IL-18 (34). In contrast, vertebrate DNA did not elicit such effects. Therefore, the larger amount of DNA administered during im vaccination may be responsible for the polarized Th1 pattern of immune response induced (32). This is due to the fact that for gene gun immunization lower doses of DNA are sufficient to induce the synthesis of immunogenic amounts of antigen, but are insufficient to trigger the production of cytokines responsible for the polarization of the T cell response.
Concluding remarks
In this study, we have compared the immune responses generated by two different delivery systems of genetic vaccination. The characteristic immune response induced by gene gun or im immunization has multiple applications in the field of infectious diseases, allergy, and cancer. The dominance of the Th1-like response induced by im injection of plasmid DNA can be used to modulate an ongoing Th2 immune response, a hallmark of asthma and allergic disorders. In contrast, the gene gun technology may be applied in the treatment of patients with autoimmune diseases who mostly show an inappropriate Th1 immune response. Nevertheless, induction of IgG2a antibodies by gene gun-mediated DNA vaccination contradicts the widely held belief that antibody responses induced by this method are restricted to those that are Th2 dependent (35). In our experiments using the gene gun, we detected a mixed Th1/Th2 immune response, as indicated by the induction of antigen-specific IgG2a and IgG1. Leitner et al. (21) observed an initial Th2-based response following gene gun immunization that switched to a Th1-like profile with activation of IFN-g-secreting T cells. Additionally, the same group reported that vaccinating mice with the circumsporozoite protein-encoded gene either by gene gun or by im injection reduces the rates of Plasmodium berghei infection; however, the level of protection obtained was significant only in the case of gene gun immunization. In summary, we have yet a lot to learn about gene vaccine delivery systems and which type of immune responses they generate.
The successful use of DNA-based vaccines in different animal models has raised enormous interest in this powerful technology. However, little is known about the level of protective immunity induced by these vaccines against human diseases. Therefore, we must carefully analyze the results of human trials currently in progress to address this important issue that will have a tremendous impact on human health.
1. Ulmer J, Donnelly JJ, Parker SE, Rhodes GH, Felgner PL, Dwarki VJ, Gromkowski SH,
Deck RR, Dewitt CM, Friedman A, Hawe LA, Leander KR, Martinez D, Perry HC, Shiver JM,
Montgomery DL & Liu MA (1993). Heterologous protection against influenza by injection
of DNA encoding a viral protein. Science, 259: 1745-1749.
2. Wollf JA, Malone RW, Williams P, Chong W, Acasadi G, Jani A & Felgner PL (1990). Direct gene transfer into mouse muscle in vivo. Science, 247: 1465-1468.
3. Tang DC, Devit M & Johnston SA (1992). Genetic immunization is a simple method
for eliciting an immune response. Nature, 356: 152-154.
4. Donnelly JJ, Ulmer JB & Liu MA (1997). DNA vaccines. Life Sciences, 60: 163-172.
5. Cox GJ, Zamb TJ & Babiuk LA (1993). Bovine herpesvirus 1: Immune responses in
mice and cattle injected with plasmid DNA. Journal of Virology, 67: 5664-5667.
6. Boyer J, Ugen KE, Wang B, Agadjanyan M, Gilbert L, Bagarezzi ML, Chattergoon M, Frost P, Javadian A, Williams WV, Rafaeli Y, Ciccarelli RB, McCallus D, Coney L & Weiner DB (1997). Protection of chimpanzees from high-dose heterologous HIV-1 challenge by DNA vaccination. Nature Medicine, 3: 526-532.
7. Davis HL, Michel ML & Whalen RG (1993). DNA-based immunization induces continuous secretion of hepatitis B surface antigen and high levels of circulating antibody. Human Molecular Genetics, 2: 1847-1851.
8. Hoffman SL, Doolen DL, Sedegah M, Aguiar JC, Wang R, Malik A, Gramzinski RA, Weiss
WR, Hobart R, Norman JÁ, Margalith M & Hedstrom RC (1997). Strategy for development
of a pre-erythrocytic Plasmodium falciparum DNA vaccine for human use. Vaccine,
15: 842-845.
9. Lowrie DB, Silva CL, Colston MJ, Ragno S & Tascon RE (1997). Protection against
tuberculosis by plasmid DNA vaccine. Vaccine, 15: 834-838.
10. Wang B, Boyer J, Srikantan V, Ugen K, Gilbert L, Phan C, Dang K, Merva M,
Agadjanyan M, Newman M, Carrano R, McCallus D, Coney L, Williams WV & Weiner DB
(1995). Induction of humoral and cellular immune responses to the human immunodeficiency
type 1 virus in nonhuman primates by in vivo DNA inoculation. Virology, 211:
102-112.
11. Wang B, Merva M, Dang K, Ugen K, Boyer J, Williams WV & Weiner DB (1994). DNA inoculation induces protective in vivo immune responses against cellular challenge with HIV-1 antigen-expressing cells. AIDS Research Human Retroviruses, 10 (Suppl 2): S35-S41.
12. Schirmbeck R, Bohm W, Ando K, Chisari FV & Reimann J (1995). Nucleic acid
vaccination primes hepatitis B virus surface antigen-specific cytotoxic T lymphocytes in
nonresponder mice. Journal of Virology, 69: 5929-5934.
13. Walker PS, Scharton-Kersten T, Rowton ED, Hengge U, Bouloc A, Udey MC & Vogel JC (1998). Genetic immunization with glycoprotein 63 cDNA results in a helper T cell type 1 immune response and protection in a murine model of leishmaniasis. Human Gene Therapy, 9: 1899-1907.
14. Porter KR, Kochel TJ, Wu SJ, Raviprakash K, Phillips I & Hayes CG (1998). Protective efficacy of a dengue 2 DNA vaccine in mice and the effect of CpG immuno-stimulatory motifs on antibody responses. Achives of Virology, 143: 997-1003.
15. Yu Z, Karen KL, Kananget S, Manickam E & Rouse BT (1998). Protection by minigenes: a novel approach of DNA vaccines. Virology, 16: 1660-1667.
16. Fynan EF, Webster RG, Fueller DH, Haynes JR, Santoro JC & Robinson HL (1993).
DNA vaccines: protective immunizations by parenteral, mucosal, and gene-gun inoculations. Proceedings
of the National Academy of Sciences, USA, 90: 11478-11482.
17. Webster RG, Fynan EF, Santoro JC & Robison HL (1994). Protection of ferrets
against influenza challenge with a DNA vaccine to the haemagglutinin. Vaccine, 12:
1495-1498.
18. Hui KM, Sabapathy TK, Oei A & Chia TF (1994). Generation of allo-reactive
cytotoxic T lymphocytes by particle bombardment-mediated gene transfer. Journal of
Immunological Methods, 171: 147-155.
19. Fuller DH & Haynes JR (1994). A qualitative progression in HIV type 1 glycoprotein 120-specific cytotoxic cellular and humoral immune responses in mice receiving a DNA-based glycoprotein 120 vaccine. AIDS Research Human Retroviruses, 10: 1433-1441.
20. Zarozinski CC, Fynan EF, Selin LK, Robinson HL & Welsh RM (1995). Protective
CTL-dependent immunity and enhanced immunopathology in mice immunized by particle
bombardment with DNA encoding an internal virion protein. Journal of Immunology,
154: 4010-4017.
21. Leitner WW, Seguin MC, Ballou WR, Seitz JP, Schultz AM, Sheehy MJ & Lyon JA
(1997). Immune responses induced by intramuscular or gene gun injection of protective
deoxyribonucleic acid vaccines that express the circumsporozoite protein from Plasmodium
berghei malaria parasites. Journal of Immunology, 159: 6112-6119.
22. Vanderzanden L, Bray M, Fuller D, Roberts T, Custer D, Spik K, Jahrling P, Huggins
J, Schmaljohn A & Schmaljohn C (1998). DNA vaccines expressing either GP or NP genes
of Ebola virus protect mice from lethal challenge. Virology, 246: 134-144.
23. Chang SW, Bu J, Rompato G & Garmendia AE (1998). A vector DNA vaccine encoding pseudorabies virus immediate early protein demonstrates partial protection in mice against lethal virus challenge. Viral Immunology, 11: 27-36.
24. Herrmann JE, Chen SC, Fynan EF, Santoro JC, Greenberg HB, Wang S & Robinson HL
(1996). Protection against rotavirus infection by DNA vaccination. Journal of
Infectious Diseases, 174 (Suppl 1): S93-S97.
25. Lai WC, Bennett M, Johnston SA, Barry MA & Pakes SP (1995). Protection against Mycoplasma pulmonis infection by genetic vaccination. DNA and Cell Biology, 14: 643-651.
26. Barry MA & Johnston SA (1997). Biological features of genetic immunization. Vaccine,
15: 788-791.
27. Robinson HL (1997). Nucleic acid vaccines: an overview. Vaccine, 15:
785-787.
28. Nichols WW, Ledwith BJ, Manam SV & Troilo J (1995). Potential DNA vaccine integration into host cell genome. Annals of the New York Academy of Sciences, 772: 30-39.
29. Davis HL, Mancini M, Michel ML & Whalen RG (1996). DNA-mediated immunization to
hepatitis B surface antigen: longevity of primary response and effect of boost. Vaccine,
14: 910-915.
30. Abbas AK, Murphy KM & Sher A (1996). Functional diversity of helper T
lymphocytes. Nature, 383: 787-793.
31. Drazen JM, Arm JP & Austen KF (1996). Sorting out the cytokines of asthma. Journal
of Experimental Medicine, 183: 1-5.
32. Leclerc C, Deriaud E, Rojas M & Whalen RG (1997). The preferential induction of
Th1 immune response by DNA-based immunization is mediated by the immunostimulatory effect
of plasmid DNA. Cellular Immunology, 179: 97-106.
33. Tighe H, Corr M, Roman M & Raz E (1998). Gene vaccination: plasmid DNA is more than just a blueprint. Immunology Today, 19: 89-97.
34. Krieg AM, Yi AK, Schorr J & Davis HL (1998). The role of CpG dinucleotides in
DNA vaccines. Trends in Microbiology, 6: 23-27.
35. Feltquate DM, Heaney S, Webster RG & Robinson HL (1997). Different T helper
cell types and antibody isotypes generated by saline and gene gun DNA immunization. Journal
of Immunology, 158: 2278-2284.
Address for correspondence: S.C. Oliveira, Departamento de Bioquímica e Imunologia, ICB, UFMG, Caixa Postal 486, 30161-970 Belo Horizonte, MG, Brasil. Fax: +55-31-499-2666. E-mail: scozeus@mono.icb.ufmg.br
Presented at the International Symposium "The Third Revolution on Vaccines: DNA Vaccines", Belo Horizonte, MG, Brasil, November 3-7, 1997. Research supported by FAPEMIG, PADCT/CNPq, PRPq/UFMG, and CNPq. Received November 19, 1998. Accepted December 9, 1998.

![[View larger version of this image (47 K GIF file)]](3388i01.gif){kind=link}
![[View larger version of this image (6 K GIF file)]](3388i02.gif){kind=link}
![[View larger version of this image (44 K GIF file)]](3388i03.gif){kind=link}